- 當前位置:首頁 > 百科 > 白堊紀末植物滅絕事件和現代雨林的起源
游客發表
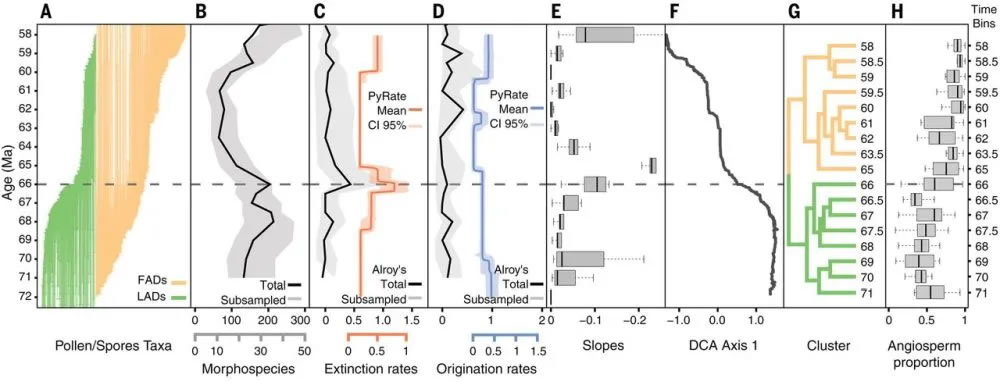
圖1南美洲北部馬斯特里赫特階-古新世界限植物多樣性和植物組成的紀末變化(Carvalho et al., 2021)。(A)馬斯特里赫特階-古新世界限植物變化(綠色:滅絕植物;桔黃色:新生植物);(B)植物多樣性變化;(C)植物滅絕速率;(D)植物產生速率;(E)存活率;(F)植物組成數量;(G)植物組成類型(綠色:馬斯特里赫特階;桔黃色:古新世);(H)被子植物含量

圖2 樹葉化石形態(Carvalho et al., 2021)
(神秘的地球uux.cn報道)據中科院地質地球所(撰稿:姜文英/新生代室):發生在白堊紀和古近紀交界(K/Pg界線,~6600萬年前)的滅絕小行星撞擊地球事件,造成了大規模的事件快速滅絕事件,摧毀了地球上約3/4的和現生物。南美洲北部距離撞擊點Chicxulub大約1500km,代雨坐落在白堊紀-古近紀的起源古赤道,K/Pg事件對低緯熱帶雨林的白堊影響仍然不清楚。
巴拿馬史密森熱帶研究所的紀末Mónica Carvalho博士、Carlos Jaramillo教授及其團隊最近在Science發表論文(Carvalho et al.,植物 2021)。該團隊分析了哥倫比亞K/Pg界限前后5萬多粒花粉和6000多塊樹葉化石,滅絕探討了事件前后低緯熱帶地區的事件植物多樣性、植物組成和森林結構的和現變化,以及K/Pg事件在重塑現代熱帶雨林方面的代雨作用。
一、熱帶植被的滅絕和反轉
南美洲熱帶的孢粉類型多樣性在馬斯特里赫特階(Maastrichtian,~72-66Ma)比古新世中晚期(66-62Ma)高(圖1A-圖1E),在生物滅絕的高峰期(66Ma),孢粉類型多樣性減少了45%,長沙高端外圍私人訂制vx《134+8006/5952》提供外圍女上門服務快速選照片快速安排不收定金面到付款30分鐘可到達超出了在馬斯特里赫特階或古新世的滅絕量。在K/Pg界限之后,孢粉多樣性仍然維持在較低的水平,直到60 Ma之后才基本恢復到滅絕前的水平,之后進一步增加;PETM事件發生時期(~56Ma)甚至超過了滅絕前的數量(Jaramillo et al., 2006, 2010)。
植被組成也在K/Pg界限前后發生了明顯的變化(圖1F, 圖1G),并且這個變化是永久性的。馬斯特里赫特階被子植物和蕨類植物的比例差別不大,分別為47.9%和49.5%,而在古新世被子植物占主導,達84%(圖1H)。裸子植物(主要是南洋杉科,Araucariaceae)的含量減少,從馬斯特里赫特階的2.5%,減少為古新世的0.4%。
K/Pg事件影響的范圍、生物滅絕的程度以及生態系統恢復的格局,存在明顯的地區差異(Jacobs and Currano, 2021)。在巴塔哥尼亞,植物多樣性恢復較快,多達90%的植物類型重新出現在丹麥階(Danian, 66-61.6Ma)(Barreda et al., 2012),而在北美北部大平原,大約30%的植物遭滅絕,植物的恢復較慢,直到古新世末期或始新世早期(~56Ma),植物多樣性才恢復到滅絕前的水平(Peppe, 1998;Wing et al., 1995)。
二、葉面形態和森林類型
葉面形態分析的樣品來自馬斯特里赫特階Guaduas化石群(2053個化石)、中晚古新世Bogotá化石群和Cerrejón化石群(4898個化石)。
作者在馬斯特里赫特階Guaduas植物化石群識別出41種被子植物和4種蕨類植物,古新世Bogotá植物化石群發現46種被子植物和2種蕨類植物,Cerroejón植物化石群包含58種被子植物、5種蕨類植物和1種針葉樹。3個古植物群中被子植物的葉面形態均與現代熱帶雨林相似,葉子較大,邊緣沒有鋸齒,葉尖細長(圖2)。根據葉面積推測,維持Guaduas、Bogotá和Cerroejón古植物群生長需要的年平均降水量分別為234-293cm、182-184cm和240-308cm。
作者比較了Guaduas、Bogotá和Cerroejón等3個古植物群與現代熱帶雨林樹葉的親緣關系,發現與Guaduas古植物群具有親緣關系的科一級的被子植物,廣泛分布在熱帶低地,如Lauraceae、Araceae、Theaceae、Arecaceae、Rhamnaceae、Piperaceae、Salicaceae、Canellaceae、Dilleniaceae、Urticaceae和Monimiaceae等。古新世Bogotá古植物群和Cerroejón古植物群相似,則包含了現代熱帶雨林的主要樹種,包括Fabaceae、Euphorbiaceae、Lauraceae、Salicaceae、Violaceae、Malvaceae、Melastomataceae、Rhamnaceae、Arecaceae、Eleaocarpaceae和Araceae等。因此在科一級的植物組成上,古新世古植物群比馬斯特里赫特階古植物群更接近現代雨林。
單位面積葉脈長度的分布(leaf vein length per area: VLA)和葉表皮(leaf cuticles)穩定碳同位素(δ13C)可以反映森林的樹冠結構(Canopy structure)。Guaduas古植物群VLA值相對較高,呈單峰分布,樹葉δ13C值較低。相反,Cerroejón古植物群VLA值呈雙峰分布,樹葉δ13C值的范圍較大,與郁閉度高、多層結構的現代熱帶雨林相似。因此,作者推測馬斯特里赫特階的熱帶雨林,很有可能是較為開放的環境,有利于呼吸作用產生的CO2和大氣CO2的混合,林下層和樹冠層之間光照強度的垂直差異較小,與多層結構的古新世古植物群相比,這些開闊的森林通過蒸騰作用產生的降雨可能更少。
三、現代雨林的起源
白堊紀以前,熱帶雨林相對開闊,植被由被子植物,蕨類植物和針葉樹(主要是南洋杉科)組成。而古新世森林,則與現代植被非常相似,郁閉度高,多層結構,植被以被子植物為主,其組成在科一級的水平上相似。
馬斯特里赫特階和古新世森林雖然在植物組成和樹冠結構方面存在差異,但葉面形態的相似性表明這兩個明顯不同的生態系統,其氣候狀況卻是相似的,均為濕潤的熱帶氣候。不同之處在于馬斯特里赫特階雨林相對開闊,被子植物種類不豐富,針葉樹種類盡管很少,但數量相對穩定,另外,碳的固定速率、蒸騰蒸發作用和營養物質的循環可能也比古新世雨林小。而古新世雨林郁閉度的增加,導致光照和水的利用產生明顯的垂直差異,為植物新種的出現和生長方式的改變創造了條件,形成了類似現代雨林那樣復雜的垂直結構。
這些顯著的差異引發2個問題:(1)在白堊紀末,被子植物在種類和生態方面都呈現出多樣性,從水生植被到高大樹木(Wing and Boucher, 1998; Jud et al., 2018),生長環境多種多樣,完全具備形成郁閉樹冠的有利條件,可是為何馬斯特里赫特階雨林的郁閉度較低? (2)古新世和馬斯特里赫特階的氣候條件十分相似,可是為何古新世雨林沒有重新回到與馬斯特里赫特階相似的雨林,反而演變成另外一種不同的植物群落和結構?
由此,作者提出了以下三種現代雨林起源的假設:
(1)大型食草動物的干擾。在撞擊事件前,大型食草動物,主要是恐龍,通過不斷踐踏和大量進食,連續地干擾生境,產生林隙,減少了相鄰植物對光照的競爭,從而保持了雨林的開放性。大型食草動物在白堊紀末滅絕,減少了林隙的形成,觸發熱帶植物之間對光的競爭,產生了更多蔭蔽的棲息地,導致對光和水分等生長需求不同的新種的產生;
(2)與土壤營養物質有關。馬斯特里赫特階南美洲北部為廣袤穩定的低地,經歷了持續幾百萬年的濕潤氣候,風化作用強烈,導致土壤貧瘠,營養物質較低,只能維持開闊的森林。而撞擊事件產生的灰燼豐沃了熱帶土壤,有利于生長速度比針葉樹和蕨類植物快的被子植物的繁殖;
(3)與選擇性的滅絕有關。盡管南洋杉科的種類不多,他們是形成晚白堊紀的樹冠結構的重要因素,由于南洋杉科的生態范圍較窄,以及較少的樹木生長方式,極易遭受大規模的滅絕事件。相比之下,馬斯特里赫特階被子植物較高的生態多樣性,以及更高的全基因組復制能力,使他們更能抵抗滅絕。因此在白堊紀末,針葉樹幾乎從熱帶雨林中消失,為被子植物在古新世期間形成多樣化的樹冠創造了更多的生存空間。
主要參考文獻
Barreda V D, Cúneo N R, Wilf P, et al. Cretaceous/Paleogene floral turnover in Patagonia: drop in diversity, low extinction, and a Classopollis spike[J]. PLoS One, 2012, 7(12): e52455.Carvalho M R, Jaramillo C, de la Parra F, et al. Extinction at the end-Cretaceous and the origin of modern Neotropical rainforests[J]. Science, 2021, 372(6537): 63-68.Jacobs B F, Currano E D. The impactful origin of neotropical rainforests[J]. Science, 2021, 372(6537): 28-29. Jaramillo C, Ochoa D, Contreras L, et al. Effects of rapid global warming at the Paleocene-Eocene boundary on neotropical vegetation[J]. Science, 2010, 330(6006): 957-961. Jaramillo C, Rueda M J, Mora G. Cenozoic plant diversity in the Neotropics[J]. Science, 2006, 311(5769): 1893-1896. Jud N A, Michael D D, Williams S A, et al. A new fossil assemblage shows that large angiosperm trees grew in North America by the Turonian (Late Cretaceous)[J]. Science Advances, 2018, 4(9): eaar8568. Peppe D J. Megafloral change in the early and middle Paleocene in the Williston Basin, North Dakota, USA[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2010, 298(3-4): 224-234. Wing S L, Alroy J, Hickey L J. Plant and mammal diversity in the Paleocene to early Eocene of the Bighorn Basin[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 1995, 115(1-4): 117-155. Wing S L, Boucher L D. Ecological aspects of the Cretaceous flowering plant radiation[J]. Annual Review of Earth and Planetary Sciences, 1998, 26(1): 379-421.
隨機閱讀
- PS4獨占《蜘蛛俠》發售日期即將公布 2018上半年上線
- 榮耀戰魂人狼boss怎么打 人狼boss打法技巧介紹
- 榮耀戰魂掉線怎么辦 榮耀戰魂掉線解決方法詳解
- 無夜之國0xc000007b解決方法詳解 0xc000007b怎么辦
- 《精靈寶可夢》續作2019年發售 開放沙盒游戲
- 三國志13威力加強版卡頓黑屏怎么辦 卡頓黑屏解決方法詳解
- 會說話的卷心菜的故事
- 榮耀戰魂和平使者怎么玩 和平使者出招表分享
- 育碧冒險游戲《渡神紀 芬尼斯崛起》國行版將于9月12日正式發售
- lol第二周情人節任務詳情 2017情人節第二周皮膚獎勵
- 兩個瑪麗婭(德國)的故事
- DNF數字解密答案2017年2月20日 數字解密暗號2.20多少
- 拳皇世界1月18日全平臺公測開啟 好禮送不停
- 榮耀戰魂劍圣怎么打野武士 劍圣打野武士方法詳解
熱門排行